|
|
 发表于 昨天 13:19
|
发表于 昨天 13:19
|
查看: 4 |
回复: 0
原文:Canadian Journal of Zoology. 2026, 104: 1-15. https://doi.org/10.1139/cjz-2025-0074
作者:S.M. Jackson, M.P. Miller, and S. Lingle
原标题:Pest or pest control? Coyote interactions with cattle and Richardson's ground squirrels
【以下均为机翻,仅供参考。】
摘要:
郊狼(Canis latrans)在加拿大草原上无处不在,每年因伤害或杀死牛(Bos taurus)而造成的损失高达数百万美元。深入了解郊狼在牛群附近的行为或许能减少这些损失。我们假设郊狼能从牛群牧场获得直接(如杀死幼牛或觅食牛群)和间接(如捕猎本地猎物)获得猎物,再进一步假设奶牛会对郊狼做出防御性反应。5月至8月,我们在萨斯喀彻温省西南部对郊狼进行了58次焦点观察,并记录了活动情况、猎物种群、与牛群的距离以及牛群对郊狼的反应。郊狼捕猎本地猎物的频率高于与牛群互动的频率。在捕猎它们最常见的猎物——理查森地松鼠(Urocitellus richardsonii,原称Spermophilus richardsonii (Sabine, 1822))时,郊狼经常出现在距离牛群100米以内的地方。郊狼会从牛尸上觅食(N=5),偶尔接近幼牛(N=2),驱赶牛群(N=2),或吃掉胎衣(N=1)。在43%的观察事件中,当牛和/或幼牛在10米范围内遭遇郊狼时,会追赶它们。我们的数据表明,郊狼主要利用牛群牧场来捕猎本地猎物,尽管偶尔有机会从牛群身上觅食或杀死幼牛,也可能促使它们利用这些区域。
前言:
以灰狼(Canis lupus)为代表的大型捕食者对牲畜构成的捕食威胁,长期以来一直是人们将这些捕食者视为“西部之敌”这一观念形成的重要因素(Jones 2002)。牲畜损失每年给生产者造成数千美元的经济负担(Muhly and Musiani 2009)。在加拿大,牛群存栏量最大的地区集中在艾伯塔省和萨斯喀彻温省(Statistics Canada 2022);这两个省份的南部地区曾主要由原生草原覆盖(Atkinson 2009; Elofson 2012)。在这些地区,美洲野牛(Bos bison)、叉角羚(Antilocapra americana)和麋鹿(Cervus canadensis)等原生食草动物,已在很大程度上被家牛(Bos taurus)所取代。狼和灰熊(Ursus arctos)等大型捕食者大多已被区域性灭绝(Atkinson 2009);这使得郊狼(Canis latrans)得以跃升为该生态系统中的顶级捕食者,这一现象很可能属于典型的“中级捕食者释放效应”(Prugh et al. 2009; Ripple et al. 2013)。与其他中级捕食者一样,郊狼具有高度多样化的食性(Prugh et al. 2009)。通过向亲代习得的适应性行为,郊狼对人类的容忍度往往会随之提高(Schell et al. 2018)。正是凭借这些特性,郊狼及其他中级捕食者得以在人类聚居区附近繁衍生息,但这同时也增加了它们与人类发生负面互动的风险(Prugh et al. 2009)。事实上,郊狼长期以来一直被视为对牲畜构成“祸害”的物种(Fitch 1948),并被认为导致了严重的牲畜损失(Canadian Cattlemen Staff 1949, reprinted 2015)。对萨斯喀彻温省作物保险公司(SCIC)2011至2012财年的分析显示,在所有因捕食造成的索赔案件中,郊狼涉案比例高达86%;而在针对郊狼捕食行为提出的牲畜赔偿索赔中,涉及牛只的索赔占到了71%(Gauthier 2016)。当年,SCIC为涉及郊狼与牲畜冲突的索赔案件支付了超过150万美元(Gauthier 2016)。而在2022至2023财年,SCIC支付的捕食赔偿金已超过300万美元(SCIC 2023)。尽管在预防工作上的支出有所增加,但2022至2023财年收到的捕食索赔案件数量仍高于此前连续五个财年的水平。
然而,鉴于郊狼具有拾荒食腐的习性,它们也被人们称为“大自然的清道夫”(Bradford 2019)。牲畜尸体(腐肉)有可能为捕食者种群提供食物补给;这种补给既可以起到缓冲作用,减少捕食者对本地猎物的捕食压力(Ciucci et al. 2020),也可能通过“表观竞争”效应(apparent competition)加剧捕食者对本地猎物的捕食行为(Holt 1977)——这其中也包括那些处于濒危边缘的物种,而在萨斯喀彻温省西南部地区,此类濒危物种的数量相当可观(Martin et al. 2018)。那些能够接触到牛只尸骨堆(boneyards)的狼群会定期前往觅食(Morehouse and Boyce 2011; Petroelje et al. 2019),其日常食谱中近四分之一的食物均来源于这些拾荒得来的牛只尸体(Petroelje et al. 2019)。相比那些无法接触到尸骨堆的狼群,依赖尸骨堆拾荒的狼群其活动迁徙范围明显较小;这一现象揭示了牲畜尸体这一“食物补给源”确实有能力对捕食者的饮食结构及其行为模式产生影响(Petroelje et al. 2019)。Ciucci等人(2020)指出,若牲畜尸体这一食物补给来源十分充裕,将会导致狼群捕食本地猎物的数量相应减少,而这一变化极有可能对其作为“顶级捕食者”所应发挥的生态角色产生深远影响。牛尸堆积场能吸引20多公里以外的郊狼前来(Kamler et al. 2004),这可能会增加郊狼在牧场区域的活动频率,进而导致其滋扰行为的增加(Goodale et al. 2015)。许多牧场主往往采取致死性手段来应对这些滋扰行为及捕食侵害。尽管有选择地移除繁殖配偶能在短期内减少损失,但若食物来源依然充裕,新的郊狼往往会迁入那些原有同类已被捕杀的区域(Blejwas et al. 2002),从而限制了这种策略的有效性。
牛群的特征(例如幼牛的年龄)可能会影响其遭受捕食的风险。土狼对牛群的捕食行为,大多集中在春季针对刚出生的幼牛(Boggess et al. 1978)。如果母牛因分娩并发症而身体虚弱,其刚出生的幼牛可能会显得尤为脆弱(Gilliland 1995)。夏季期间,土狼对牛群的捕食行为往往呈下降趋势(Boggess et al. 1978);这可能归因于幼牛躲避捕获的能力随时间增强(Green 1994)。土狼往往偏好在有家养或野生有蹄类动物幼崽出没的区域进行捕猎(Paquet 1992; Blejwas et al. 2002),但牲畜所表现出的防御行为能够有效震慑土狼的攻击(Connolly et al. 1976)。当母牛为保护幼牛免受感知到的威胁而发起防御时,其攻击行为甚至可能造成致命后果(Turner and Lawrence 2006);此外,母牛之间为共同保护幼牛而采取的协同防御策略,可能是部分牧场主之所以愿意容忍捕食者存在的一个重要因素(Allen and Fleming 2004)。然而,鉴于牛群各项性状的可遗传性因品种而异(Titterington et al. 2022),因此不排除某些个体(Flörcke et al. 2012)、特定种群或特定品种的母牛,在保护幼牛方面可能表现得不如其他牛群那样积极。例如,海福特牛(Hereford)和安格斯牛(Angus)就被描述为性情尤为温顺的品种(Hoppe et al. 2010);这类性格特质可能会影响动物抵御捕食者侵害(无论是保护自身还是其后代)的能力(Réale and Festa-Bianchet 2003)。
郊狼在捕猎体型较大的猎物时,通常会采取群体协作的方式(Gese et al. 1988; Lingle 2000)。野生有蹄类动物虽然是土狼日常食谱中的常规组成部分,且主要集中在冬季被大量捕食(Bekoff and Wells 1980; Bowen 1981; Lingle 2000; Lingle et al. 2022),但其活动范围往往与牛群的放牧区域存在重叠(Yoakum 1975; Cooper et al. 2008)。郊狼通常更倾向于单独或成对活动,以便独自捕猎较小的猎物(Lingle 2000);它们也常在牛群放牧的牧场上捕食小型哺乳动物,且通常与牛群相安无事(Fitch 1948)。在草原地带,从春季到秋季,郊狼普遍以理查森地松鼠(Urocitellus richardsonii (Sabine, 1822),旧称Spermophilus richardsonii (Sabine, 1822))及其他松鼠科动物为食(Lingle 2000; Lingle et al. 2022)。
郊狼在牛群牧场上捕猎理查森地松鼠之所以能取得更高的成功率,可能归因于以下几个原因。首先,理查森地松鼠往往更集中地分布在牲畜利用的原生牧场以及植被低矮的区域(Downey et al. 2006; Bylo et al. 2014),这种分布特征增加了郊狼与该猎物物种相遇的几率,从而提高了捕猎成功率。其次,牛群牧场上低矮的植被改善了郊狼侦测或捕捉地松鼠的条件(Wells and Bekoff 1982),这也是郊狼在此类牧场捕猎地松鼠更为得力的原因之一。第三,牛群的存在可能对郊狼有利,因为牛群本身可能会阻碍地松鼠察觉或听见正在逼近的郊狼。若情况确实如此,我们便可推测:在一天中地松鼠最为活跃的时段,郊狼往往会选择靠近牛群活动。
萨斯喀彻温省西南部地区呈现出一种由林地牧场与放牧牧场交织而成的景观格局,这使得当地的牧场主在草原生态管理中扮演着至关重要的角色(Atkinson 2009)。既往研究报告显示,在萨斯喀彻温省西南部部分地区,牛类在郊狼的食谱中占比近半(Lingle et al. 2022);据当地牧场主的经验判断,郊狼所食用的牛类主要来源于牛尸(腐肉)。然而,该地区有关郊狼捕杀牲畜(即“掠食”)的报告也屡见不鲜(Gauthier 2016)。深入了解郊狼如何利用牛群牧场以及它们与牛群之间如何互动,不仅有助于影响牧场主对郊狼的容忍态度,还能为建立有效的牲畜防掠食机制奠定坚实的基础。
在本项研究中,我们针对郊狼在牛群牧场上的活动特征,检验了三项彼此之间并非相互排斥的假设。针对假设1,我们检验了郊狼是否因从牛群中获取直接利益(例如捕杀幼牛或取食牛尸)而频繁利用牛牧场。我们预测,在牛牧场活动的郊狼会捕猎(接近或攻击)或杀死幼牛,骚扰母牛以获取胎盘等副产品,并取食牛尸。针对假设2,我们检验了郊狼是否通过除将牛作为食物来源以外的其他方式,从牛牧场及其与牛群的近距离接触中获益。我们预测,在牛牧场活动的郊狼捕猎本地猎物物种的频率高于捕猎牛的频率,且它们停留在牛群附近(<100米)的时间多于远离牛群的时间。我们进一步预测,在牛牧场活动的郊狼,当处于牛群附近(<100米)时,比处于远离牛群的位置时,更有可能捕猎理查森地松鼠,且捕猎成功率更高。最后,针对假设3,我们检验了该地区的母牛是否会对郊狼的出现做出防御性反应。我们预测,当郊狼遭遇牛群时——特别是在幼牛较易遭受捕食的时期——母牛会表现出防御行为(如聚拢成群或驱逐郊狼);反之,则说明母牛缺乏反捕食防御能力,可能反映了牛在驯化过程中对捕食者的反应有所松弛。
研究方法:
研究地点
本研究于2022年5月16日至8月7日在大草原国家公园(GNP)生态系统内开展。研究地点涵盖了延伸至GNP西区以北约27.5公里处的牛牧场,以及位于GNP西区(面积约525平方公里)内部、可俯瞰田野与河谷的丘陵地带(其中部分区域亦有牛群分布);整个研究区域总面积约为2600平方公里。大GNP生态系统地势起伏显著,主要地形特征包括连绵起伏的丘陵和陡峭的河谷。该区域的植被主要由原生混合草草原构成(加拿大公园管理局 2018),但也包含部分耕地(图1)。在2022年的观测季期间,平均气温介于5月份的9.9 °C与8月份的20.9 °C之间。
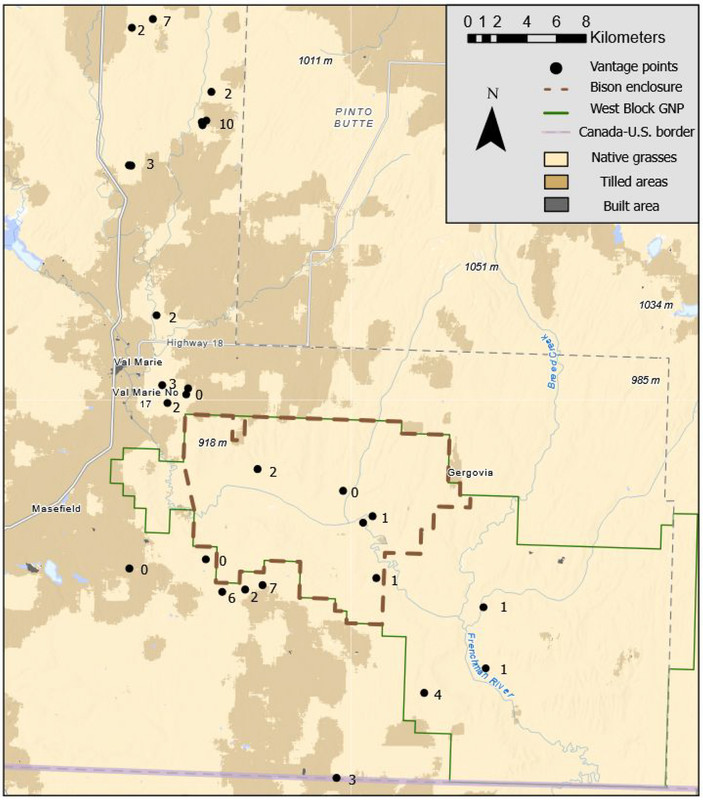
图1. 研究地点地图,展示了用于观察郊狼(Canis latrans)的观测点、美洲野牛(Bos bison)围栏(该围栏将家牛排除在外)、“西区”(West Block)边界、原生草地、耕作区以及建筑区。每个观测点(或一组观测点)旁标注的数字,表示在该位置针对郊狼成功开展的焦点观察次数。数值为0表示我们未能从该位置成功开展针对郊狼的焦点观察。本图使用 ArcGIS Pro (ESRI) 制作,数据信息来源于 Karra 等人 (2021) 和加拿大公园管理局 (2021)。底图服务图层致谢:Esri Canada, Esri, TomTom, Garmin, SafeGraph, METI/NASA, USGS, EPA, NPS, USDA, NRCan, Parks Canada, Esri, NASA, NGA, USGS, Impact Observatory, Microsoft, 以及 Esri。地图投影:NAD 1983;坐标系:UTM Zone 13N。
大GNP生态系统(Greater GNP Ecosystem)为许多受威胁物种提供了关键栖息地,其中包括大艾草鸡(Centrocercus urophasianus)、穴鸮(Athene cunicularia)及其他几种地栖鸟类、黑尾草原犬鼠(Cynomys ludovicianus)和敏狐(Vulpes velox)(Parks Canada Agency 2016, 2021)。黑尾鹿(Odocoileus hemionus)和白尾鹿(Odocoileus virginianus)栖息于该国家公园及其周边地区,且在秋季至早春期间常被郊狼捕食(Lingle et al. 2022)。此外,驼鹿(Alces alces)和叉角羚也分布于此。
牛群在国家公园内外均有放牧(图1)。围栏虽能阻隔牛群和大角野牛的移动,却无法拦住郊狼、麋鹿、驼鹿、鹿类、叉角羚及地松鼠等体型较小的动物。牧场主通常会在12月前后将牛群赶至靠近宅基地(距离建筑物数公里范围内)的地方,因为冬季期间牧场主需对牛群进行人工补饲;随后在4月或5月(通常是在大多数牛犊出生之后),他们会将母牛及其牛犊组成的牛群转移至距离宅基地较远的牧场进行放牧。在开展野外考察工作之前,我们曾与牧场主进行座谈,以了解不同季节牛群的具体位置,并听取他们关于郊狼的经历与看法。我们依据从这些座谈中获取的信息,确定了用于观察郊狼的最佳观测点(图1)。
在萨斯喀彻温省(Saskatchewan),猎杀郊狼无需申领狩猎许可证,且猎人可将猎获的郊狼尸体上交至当地乡村市镇政府以换取经济补偿;此外,该省对郊狼的狩猎活动不设禁猎期(Saskatchewan Wildlife Regulations 1981)。部分当地居民一旦发现郊狼便会将其射杀;而另一些居民出于多种原因,对郊狼持较为友善的态度——例如,他们认为郊狼在清理可能传播疾病的动物尸体方面发挥着积极作用。
物种鉴定
Green (1994) 将郊狼描述为一种犬科动物,成年个体的体重估计在 10–16 千克之间,特征包括“直立的尖耳、细长的吻部以及浓密的尾巴”。郊狼可通过体长和耳长等形态特征与狼区分开来(Bekoff 1977; Wheeldon and Patterson 2012);不过,狼在该研究区域内已处于区域性灭绝状态(Mech 1974; Atkinson 2009)。红狐(Vulpes vulpes)和敏狐均分布于该研究区域内;其中,红狐具有显眼的白尖尾巴(Lariviѐre and Pasitschniak-Arts 1996),而敏狐则具有黑尖尾巴(Egoscue 1979),但这两种狐狸的体型均小于郊狼。
成年理查森地松鼠(Richardson's ground squirrels)的体重约为 200–600 克,具体数值取决于性别和季节等因素(Michener 1984, 1998)。它们可通过其浅黄褐色的体色和尖锐的鸣叫声与其他地松鼠区分开来(Michener and Koeppl 1985)。黑尾草原犬鼠的体型较大,且尾巴末端呈黑色(Hoogland 1996);相比之下,十三线地松鼠(Ictidomys tridecemlineatus (Mitchill, 1821),旧称 Spermophilus tridecemlineatus (Mitchill, 1821))体型较小,且背部具有深色条纹(Streubel and Fitzgerald 1978)。
尽管区分家牛品种可能较为困难,且若要获得确切结果通常需要进行分子分类鉴定,但在实际应用中,地理分布以及体色、角形和颅骨形态等表型特征仍是常用的鉴定依据(Felius et al. 2011)。基于牛只的体色特征以及牧场主提供的信息,我们将研究区域内的雌性及幼年牛只鉴定为以下品种之一:安格斯牛(黑色安格斯,通体黑色)、红安格斯牛(通体深红色)、海福特牛(棕白相间),或是杂交牛(即上述两个品种的杂交后代,兼具两者的特征)。
观察方法
从2022年5月中旬至8月上旬,两名观察员(SMJ和MPM)共耗时158小时,蹲守在俯瞰田野和山谷的山丘上,观察郊狼与牛群及本地猎物物种之间的互动。我们使用高倍双筒望远镜和单筒望远镜来观察1000至2000米以外的动物,以确保不对其造成惊扰。我们共进行了62次观察,平均每次持续154 ± 7分钟(平均值 ± 标准误)。在这62次观察中,有50次我们至少对一只郊狼进行了一次“焦点观察”(focal observation),最终共计完成了58次针对特定郊狼的观察会话。被选为焦点的郊狼平均处于视野内的时间为35 ± 5分钟。我们每天进行两次观察:第一次从日出前约1小时开始,持续至日出后3至4小时;第二次从日落前3至4小时开始,持续至日落时分。如果在观察开始后的2小时内未能发现郊狼,我们会随即结束该次观察。否则,我们会在观察持续满4小时后结束;或者在最后一次目击郊狼的1小时后结束;抑或当光线变得过于昏暗、无法继续观察动物时结束。
我们将每次观察期间首次被发现的郊狼选定为“焦点郊狼”。如果同时发现多只郊狼结伴成群(定义为某只郊狼与至少另一只郊狼的距离在100米以内,见表1),则选取该群体中始终走在最前方的郊狼作为焦点。如果同时发现两群或多群郊狼,且它们行进的方向各不相同,我们会选择观察其中一只似乎正向着某一特定区域移动的郊狼——即预计它在该区域内能被持续观察到最长时间的那只郊狼。如果某只郊狼脱离视野超过30分钟,且此时视野内尚有其他郊狼可见,我们会随即开始对另一只郊狼进行新的焦点观察。每次观察期间包含1至5次(平均1.18 ± 0.09次)焦点观察。为了估算动物之间的距离,我们利用可见动物的大致体长(成年母牛约 2 米,鹿约 1 米,郊狼约 0.5 米),或相邻栅栏柱之间的距离(约 2 米)作为参照,进而计算该未知距离内可容纳多少只动物或多少根栅栏柱。

在观察期间,我们收集了关于作为观察焦点的郊狼个体的相关数据。在每一分钟的起始时刻,我们会进行一次瞬时扫描,以记录该郊狼的活动类型(表2)、群体规模以及与牛群之间的距离。我们将该焦点郊狼与牛群的距离划分为四个类别:距离牛群不足10米、不足100米、超过100米;或者牛群处于观察者的视线之外(不可见),抑或距离超过1000米且据推测不在该焦点郊狼的视线范围内(表1)。如果在任意1分钟的间隔内,该焦点郊狼正在进行捕猎或进食,我们会采用“一零取样法”(Martin和Bateson 2007)来记录其捕获的猎物或食物种类(若已知),以及捕猎行为是否存在任何升级。我们将“捕猎行为”的界定范围严格限制在郊狼已发现并正与处于视线内的特定猎物个体或群体进行直接互动之时;对于郊狼可能正在搜寻猎物但猎物尚未进入视线的情况,我们不将其归入捕猎行为之列。捕猎互动过程可能会逐步升级为接近、追逐、攻击或捕获等阶段(表2)。作为一种常见的追逐形式,郊狼在面对单个地松鼠时,通常会先进行短距离的猛冲,随后通过“头部猛刺”动作对猎物发起攻击(Wells和Bekoff 1982; Gese et al. 1996)。在较远的观察距离下,当郊狼针对地松鼠发生追逐(猛冲)或捕获行为时,我们能够对其进行识别并记录;但由于无法准确辨识“头部猛刺”这一细节动作,因此在记录郊狼与地松鼠的互动时,我们未将“攻击”阶段与“追逐”阶段进行细致区分。此外,郊狼也会对有蹄类动物群体发起短距离的猛冲(Gese和Grothe 1995; Lingle和Pellis 2002),其中包括牛群(详见“结果”部分);此类猛冲行为既可能属于捕食尝试的一部分,也可能旨在试探该群体中特定个体的脆弱程度。为了确保描述的清晰度,并避免对郊狼行为的意图进行主观臆测,凡涉及郊狼与牛群之间发生的接近或追逐行为,我们均仅对其具体的行为形式(例如:猛冲、步行接近等)进行客观描述。

当从观察点能够看到牛群时,我们记录了每群牛(即在同一围栏区域内、彼此可自由接触的成年母牛及其幼牛)的数量,并记录了其中是否有幼牛存在(表1)。为了考察牛群的防御行为,我们记录了牛群是否出现聚拢现象,以及是否有成年母牛或幼牛向作为观察焦点的郊狼靠近或对其进行驱赶;这里的“靠近或驱赶”被定义为一种不经停顿、且速度快于慢走的直接移动行为(例如快走、小跑、疾驰或全速奔跑),旨在逼近郊狼(Hildebrand 1965, 1977;表1)。此外,我们还记录了每个观察时段的具体日期、时间以及观察点的位置。
为了在特定的观察时段内追踪个体郊狼,一名观察员负责持续监视作为焦点的郊狼,而另一名观察员则负责监视其他的郊狼——包括与焦点郊狼同属一群的郊狼,以及属于其他群体或单独活动的郊狼。当郊狼因地形、植被、牛群等因素而从视野中消失时,我们会持续监视其可能重新显露身形的潜在位置。由于无法跨越不同日期对个体郊狼进行辨识,为了降低在不同日期重复观察到同一只郊狼的可能性,我们的观察工作被分散在约500平方公里的广阔区域内;且在任何单一的观察点,进行的焦点观察次数均未超过10次(平均观察次数为3.3次)。基于在研究区域内收集到的粪便样本所进行的分子数据分析(旨在通过基因分型识别个体郊狼)结果显示:在166份已完成基因分型的粪便样本中,有108份是由不同的个体郊狼所排泄的;其中,有38只个体曾排泄过2至5份粪便(Jackson和Lingle,未发表数据)。这些分子数据表明,该地区的郊狼种群规模较大;即便在同一地点反复收集粪便样本,重复采样到同一只个体的几率依然较低。所有的观察工作均严格遵循了《加拿大动物护理委员会指南》(温尼伯大学动物护理委员会批准文号:AEW001)的相关规定。在开展本文所述的野外工作期间,研究人员持有萨斯喀彻温省政府颁发的《学术研究许可》(编号:21AR028w),以及加拿大公园管理局颁发的《研究与采集许可》(编号:GRA-2021-41018)。
数据分析
牛群带来的直接益处
为了验证我们的首个假设——即郊狼频繁利用牛群牧场是因为它们在此捕杀幼牛或取食牛尸——我们首先考察了郊狼在牛群牧场上的活动类型(包括移动、休息或捕食/进食),随后分析了郊狼捕杀、与之互动或取食的猎物种类。鉴于数据分布不符合参数检验的前提假设,我们采用了非参数检验方法。具体而言,我们利用 R 语言基础包 `stats` 中的 Wilcoxon 符号秩检验,并结合 `effectsize` 包(Ben-Shachar 等,2020)提供的秩二列相关系数,对郊狼在移动、休息及捕食/进食这三类活动上所花费的时间进行了比较;分析对象仅限于那些在焦点观察期间至少从事过上述三类活动之一的郊狼个体。随后,我们采用 Holm–Bonferroni 方法(Holm,1979)对 *p* 值进行了校正,以控制后续三组两两比较检验中可能出现的 I 类错误(弃真错误)。在进行此项分析时,我们将焦点观察数据的筛选范围限定为持续时间超过 20 分钟的记录,旨在确保郊狼有充足的机会在观察期间从事不止一种类型的活动。针对每一类活动,我们均报告了参与该活动的郊狼群体的规模。鉴于在单次观察过程中群体规模可能发生变化,我们针对每一类活动,基于单次焦点观察期间记录到的最大群体规模,计算并报告了其平均值(±标准误差)。
此外,我们运用符号检验(Sign-test)来验证郊狼是否更倾向于捕食本地野生猎物,而非与牛群发生互动(例如接近幼牛或冲向牛群);此项检验使用了BSDA包(Arnholt 和 Evans, 2023)中的符号检验功能,并辅以rcompanion包(Mangiafico, 2023)计算的 Cliff's Delta 效应量指标。之所以选用符号检验并采用“存在/缺失”二元数据进行分析(而非直接比较各类活动所耗费的时间),主要原因在于各次观察记录的持续时长存在差异,且在所有的焦点观察记录中,均未出现郊狼既捕食本地野生猎物又与牛群发生互动的情况。最后,我们详细报告了郊狼与牛群遭遇时各类具体结果的发生频率,其中包括郊狼接近幼牛、冲向牛群、为争夺胎衣而骚扰母牛,以及郊狼取食牛尸的发生频率。由于牧场主告知了我们牛尸的存在,因此,与我们在缺乏牛尸先验知识的情况下观察郊狼的情形相比,郊狼利用尸体的频率相对于其他活动的频率,很可能显得偏高。我们仅在牧场主所指明的地点,观察到了郊狼取食牛尸的现象。
牛群带来的间接益处
为了验证我们的第二个假设——即郊狼从牛群牧场中获益,且通过紧邻牛群也能获益(其方式并非仅限于将牛群作为食物来源)——我们考察了郊狼捕食理查森地松鼠的具体时段,以及郊狼处于牛群附近的具体时段。我们采用了一个包含二项式响应变量的广义线性混合模型(GLMM),以此来确定郊狼捕食地松鼠的具体时段;在该模型中,我们将郊狼的个体编号(ID)设为随机因子,以控制重复抽样带来的影响(每分钟均记录一次目标郊狼的行为数据)。该模型及后续的GLMM均利用 lme4 软件包(Bates et al. 2015)进行拟合,并利用 car 软件包(Fox and Weisberg 2019)计算 Wald-χ² 统计量。我们将时段划分为“晨昏时段”(日出或日落前后1小时内)或“昼行时段”(日出后1至3小时内,或日落前1至3小时内),并按如下形式构建模型:(捕食理查森地松鼠) ∼ (具体时段) + (1|Coyote ID)。此外,我们利用 DHARMa 软件包(Hartig 2024)对模型残差进行了模拟与评估。QQ图和均匀性检验结果均表明,该模型符合相应的假设条件。尽管“残差-拟合值”散点图呈现出轻微的“漏斗状”分布特征,但方差齐性 Levene 检验的结果并不显著,这表明这种漏斗状分布现象尚不严重,不足以违背模型的基本假设。对随机效应(即郊狼个体编号)各水平下的 Pearson 残差进行检查后发现,其数值普遍接近于0,仅有极个别个体呈现出离群值特征。最后,我们采用符号检验(Arnholt and Evans 2023; Mangiafico 2023)来判定:当牛群与郊狼同时出现在同一区域时,郊狼是倾向于在牛群附近(距离 < 100米)停留的时间更长,还是倾向于在远离牛群(距离 > 100米)的地方停留时间更长。由于我们在日出前及日落后时段所采集的数据量不足,因此无法针对晨昏时段开展类似的分析。
我们采用了三个包含二项式响应变量的广义线性混合模型(GLMM),旨在确定:在焦点观察期间,当郊狼处于牛群100米范围内时,其捕猎或追逐地松鼠的几率,是否高于其距离牛群超过100米、或在牛群牧场上捕猎但视线内无牛群(或牛群距离超过1000米)的情况。我们将被观察的特定郊狼个体(即“焦点郊狼”)指定为随机因子,以控制在同一焦点观察时段内,对某些特定郊狼在不同距离处进行重复采样的影响。第一个模型纳入了郊狼距离牛群在100米以内或超过100米时的焦点观察数据,其拟合公式如下:(捕猎理查森地松鼠) ∼ (与牛群的距离) + (1|郊狼ID)。第二和第三个模型纳入了郊狼距离牛群在100米以内、或者观察者视线内无牛群(即牛群距离超过1000米,且推测不在焦点郊狼的视线范围内)时的焦点观察数据。这两个模型的拟合公式分别为:(捕猎理查森地松鼠) ∼ (与牛群的距离) + (1|郊狼ID),以及 (追逐理查森地松鼠) ∼ (与牛群的距离) + (1|郊狼ID)。对于这三个模型,QQ图、均匀性检验和方差齐性检验结果均符合模型的假设条件。残差对拟合值的散点图显示,在拟合值的极值处出现了极端的残差值;鉴于预测变量和响应变量均为二元变量,出现这种情况是符合预期的。针对随机效应(即“郊狼ID”)各水平所进行的Pearson残差分析显示:对于第一个模型,Pearson残差值分布在-1至3.5的范围内;对于后两个模型,Pearson残差值则分布在-1至0或1至3的范围内。
母牛和牛犊对郊狼的反应
为了检验我们的第三个假设——即该种群中的牛会采取防御行为来保护牛犊——我们考察了牛与郊狼遭遇的频率。我们将“遭遇”定义为:至少有一头母牛或牛犊处于郊狼10米范围之内(见表1);在这些遭遇中,我们记录了母牛和/或牛犊是否表现出保护行为,此类行为具体定义为:聚拢成群、接近郊狼,或追逐郊狼。我们对牛针对郊狼的行为进行了评分,无论郊狼是主动接近牛犊还是母牛;因为郊狼经常在距离牛不到10米的范围内捕食地松鼠或进行其他活动。我们利用 R 语言基础包 `stats` 中的逻辑回归模型(并结合 `car` 包 [Fox and Weisberg 2019] 提供的 Wald-χ² 检验值),来探究母牛及牛犊追逐郊狼的行为与日期之间是否存在关联。所用数据来源于针对特定个体的焦点观察记录,且仅选取那些发生了牛群与郊狼遭遇的观察案例(其中“遭遇”的定义同前,即郊狼进入了母牛或牛犊10米范围内)。
为确保统计学上的独立性,针对每一次特定的焦点观察,我们仅提取一个数据点进行分析;即便在同一次焦点观察期间,牛群与郊狼发生了多次遭遇,或牛群对郊狼进行了多次追逐,我们也仅保留一次数据记录。对于母牛而言,我们预测其追逐郊狼的行为更可能发生在观察季的中期,即有更多年幼牛犊在场的那段时间。因此,我们将日期数值进行平方处理以构建一个二次项,并将其与作为主效应的日期变量一同纳入模型之中。对于牛犊追逐郊狼的行为,我们采用了简单的逻辑回归模型,仅将日期作为主效应纳入考量;这是基于我们的一项假设:随着牛犊的成长,其自我防御能力也会随之增强。
在上述两项分析中,日期变量均被划分为五个为期两周的时间段。此外,我们还运用了符号检验(Sign-test)方法(Arnholt and Evans 2023; Mangiafico 2023),以判定母牛追逐郊狼的倾向是否高于牛犊。在此项分析中,我们同样采用了基于特定焦点观察的“发生/未发生”(presence–absence)二元数据,且仅选取那些记录了牛群与郊狼遭遇的观察案例。所有数据分析工作均利用 R 统计软件(R Core Team 2022)完成。当 p 值小于 0.05 时,我们认定其差异具有统计学显著性。
结果
郊狼的活动概览
郊狼用于移动的时间多于用于捕猎/进食的时间(图2;Wilcoxon符号秩和检验:V = 300, padj = 0.019, rrb = 0.59 [95%置信区间 = 0.23, 0.81]);但其用于移动的时间并未多于用于休息的时间(图2;Wilcoxon符号秩和检验:V = 216, padj = 0.882, rrb = 0.14 [95%置信区间 = −0.29, 0.53]),用于休息的时间也未多于用于捕猎/进食的时间(图2;Wilcoxon符号秩和检验:V = 222, padj = 0.882, rrb = 0.17 [95%置信区间 = −0.30, 0.58])。
图2. 焦点郊狼(Canis latrans)参与不同活动的时间百分比。在比较用于狩猎/进食的时间与用于移动或休息的时间时,所有的狩猎和进食活动均被合并计算。不同的字母(a、b)表示移动、休息以及狩猎/进食这三类活动所耗费的时间之间存在显著差异;N = 27只焦点郊狼。箱体线条分别代表第25百分位数、中位数和第75百分位数。须线表示最小值和最大值(不含离群值),离群值则以独立点的形式显示。对于第25和第75百分位数均为0的活动,图中不显示箱体。
牛带来的直接与间接益处
相比于与牛发生互动,郊狼更倾向于捕食本地猎物(表3;相关样本符号检验:p = 0.003,Cliff's delta = −0.65,N = 23只目标郊狼),其中理查森地松鼠是它们最常见的猎物(表3)。在郊狼进行的19次针对本地猎物的捕食活动中,有13次(68%)是捕食理查森地松鼠;其余的捕食活动则分散在不同种类的小型猎物(例如:田鼠 [Microtus pennsylvanicus (Ord, 1815); Myodes gapperi (Vigors, 1830); Lemmiscus curtatus (Cope, 1868)]、鼠类 [Mus musculus Linnaeus, 1758; Perognathus fasciatus Wied-Neuwied, 1839; Onychomys leucogaster (Wied-Neuwied, 1841); Peromyscus maniculatus (Wagner, 1845); Zapus princeps J.A. Allen, 1893]、无脊椎动物)或本地有蹄类动物之间(表3)。郊狼在移动时的平均最大群体规模为 1.72 ± 0.13(N = 54);休息时为 1.65 ± 0.27(N = 23);捕食地松鼠时为 1.38 ± 0.14(N = 13);与牛互动时为 2.25 ± 0.95(N = 4);以及捕食本地有蹄类动物时为 1.25 ± 0.25(N = 4)。
在34次涉及捕猎、与牛群互动或进食的焦点观察中,作为观察对象的郊狼曾两次(占6%)单独接近某头小牛,两次(占6%)冲向由母牛和小牛组成的牛群,并有一次(占3%)为争夺胎盘而骚扰了一头母牛。在这些观察期间,郊狼并未攻击或捕获任何单独的母牛或小牛,但确实成功获取了胎盘。在29个已记录小牛是否在场的被观察牛群中,有97%的牛群中存在小牛。所有郊狼接近、冲向或骚扰牛群的事件均发生在6月,而在7月或8月未见此类观察记录。在5次焦点观察中,记录到了郊狼对两具已知牛尸中的一具进行食腐的行为。未曾观察到郊狼在同一时间既取食牛尸又接近小牛或冲向牛群的情况。
郊狼表现出一种非显著性的倾向:相比于晨昏时段(即日出或日落前后1小时内),它们在日出后1至3小时以及日落前的时间段内,更频繁地捕猎理查森地松鼠(图3a;采用二项式响应变量的广义线性混合模型 [GLMM] 分析结果:β ± SE = 0.77 ± 0.43,优势比 [OR] = 2.16 [OR的95%置信区间 = 0.93, 5.07],Wald-χ² = 3.17,p = 0.075,N = 47只焦点郊狼)。在对可见牛群的区域进行观察,且已知焦点郊狼与牛群之间距离的情况下,郊狼在整个白昼时段内,处于距牛群100米以内范围的概率要高于处于更远距离(>100米)的概率(图3b;配对样本符号检验:p = 0.005,Cliff's delta = 0.48,N = 30只焦点郊狼);事实上,在郊狼处于距牛群100米以内的时间里,它们处于距牛群10米以内的时间占比高达49% ± 7%(平均值 ± 标准误差)。
图3. 郊狼(Canis latrans)在一天中不同时段(相对于日出和日落时间)内,(a) 捕食地松鼠(Urocitellus richardsonii,旧称Spermophilus richardsonii)的时间百分比;以及 (b) 被观测到处于距牛群(Bos taurus)不同距离处的时间百分比。正午时段周围的空白间隔分别对应:5月约5.5小时,6月约6小时,7月约5.75小时,8月约4.5小时。(a) 图:N = 58只目标郊狼;(b) 图:N = 30只目标郊狼。
我们发现,当郊狼靠近牛群(<100 m)时,其捕猎地松鼠的几率与远离牛群(>100 m)时相比,没有显著差异(图4;二项式响应GLMM模型结果:β ± SE = −1.85 ± 1.21,OR = 0.16 [95%置信区间 = 0.01, 1.67],Wald-χ² = 2.35,p = 0.125)。当郊狼远离牛群但仍身处有牛的牧场中(>100 m)时,未发生任何追逐或捕获行为。与观察者视线中无牛,或牛群距离超过1000米且推测不在目标郊狼视线范围内的情况相比,郊狼在靠近牛群(<100 m)时表现出一种非显著性的倾向,即更频繁地捕猎地松鼠;但在靠近牛群时,其追逐地松鼠的几率并未显著增加(二项式响应GLMM模型结果:捕猎,β ± SE = −1.56 ± 0.86,OR = 0.21 [95%置信区间 = 0.04, 1.12],Wald-χ² = 3.33,p = 0.068;追逐,β ± SE = −1.20 ± 0.87,OR = 0.30 [95%置信区间 = 0.05, 1.67],Wald-χ² = 1.88,p = 0.170)。在涉及郊狼捕猎地松鼠的13次焦点观察中,目标郊狼在85%的观察中对地松鼠进行了潜行(即低头缓慢向猎物移动),在69%的观察中进行了追逐,并在23%的观察中成功捕获了地松鼠。所有涉及郊狼成功捕获地松鼠的焦点观察,均包含了先潜行后猛冲的捕猎序列。
图4. 郊狼(Canis latrans)与地松鼠(Urocitellus richardsonii,原称Spermophilus richardsonii)发生互动(狩猎、追逐、捕捉)的焦点观察百分比,该百分比取决于郊狼与牛的距离。
母牛及牛犊对郊狼的反应
在21次焦点观察中,当郊狼进入牛群10米范围内时,有9次(占43%)出现了母牛和/或牛犊追逐或逼近郊狼的情况;这21次焦点观察共计发生了33次独立的追逐/逼近事件,其中包括郊狼两次逼近牛犊的案例。在两次焦点观察中,母牛与牛犊组成的群体发生了聚拢行为:一次是因遭到一群郊狼的猛冲;另一次则是因郊狼为争夺母牛的胎衣而对母牛及牛犊进行了骚扰。母牛似乎在6月中旬至7月中旬期间更倾向于追逐郊狼(图5),但我们并未发现母牛追逐郊狼的行为与日期之间存在曲线关系(β ± SEM = −0.66 ± 0.46;Wald-χ² = 2.10,p = 0.148)。同样,我们也未发现日期与牛犊追逐郊狼的概率之间存在相关性(β ± SEM = 0.45 ± 0.40;Wald-χ² = 1.28,p = 0.258)。母牛追逐郊狼的频率并未高于牛犊(图5;配对样本符号检验:p = 1.0,Cliff's delta = −0.05)。
图5. 在与牛(Bos taurus)群相遇的郊狼(Canis latrans)焦点观察中,至少有一头牛或小牛追逐郊狼的百分比。数据按2周时段划分。N=21次焦点观察,时间为2022年5月29日至8月6日,期间至少有一次观察中郊狼距离牛群10米以内。
讨论
2022年5月至8月期间,郊狼在牛牧场及靠近牛群的区域花费了大量时间。郊狼捕食本地猎物的频率高于其与牛群互动的频率,其中理查森地松鼠是它们最常见的猎物。草原犬鼠也是郊狼的常见猎物之一(Lingle et al. 2022)。我们的观察主要集中在那些分布有理查森地松鼠但缺乏草原犬鼠的牛牧场上,这解释了为何我们在本次研究中鲜少观察到郊狼捕食草原犬鼠的现象。在少数几次案例中,郊狼曾试图接近牛犊、冲向由母牛及其幼崽组成的牛群,或对牛的尸体进行食腐。尽管我们判定郊狼用于移动的时间多于用于捕食或进食的时间,但在界定捕食行为时,我们将“搜寻猎物”的时间排除在外,这一做法可能导致我们低估了郊狼相对于其他活动所投入的实际捕食总时长。在我们判定为“移动”的时间段中,有一部分时间实际上是用于搜寻猎物的。对于郊狼而言,搜寻阶段是整个捕食过程中耗时最长的阶段,但往往难以被明确无误地识别出来(Bekoff and Wells 1986; Lingle and Pellis 2002)。
我们发现部分证据支持假设1,即郊狼频繁利用牛牧场是出于从牛群身上获取直接利益的目的。然而,郊狼似乎极少能从活牛身上直接获益;相反,它们前往牛牧场可能主要是为了获取本地猎物资源。我们发现了支持假设2的证据,即郊狼确实能从牛牧场环境中获益,且靠近牛群也能带来益处——这些益处并非单纯源于将牛群作为食物来源。郊狼在靠近牛群的区域停留的时间显著长于远离牛群的区域;此外,当身处牛群附近时,郊狼捕食地松鼠的频率呈现出一种虽未达统计学显著水平、但仍可观察到的上升趋势——相比之下,当牛群处于观察者的视线之外(或距离超过1000米,且推测已不在目标郊狼的视野范围内)时,郊狼捕食地松鼠的频率则相对较低。我们还发现了支持假设3的证据,即母牛在察觉到郊狼的存在时,会采取防御性的反应。我们频繁观察到母牛追逐郊狼的现象。在此,我们将对上述各项假设所对应的预测结果进行梳理,并将本次研究的发现与此前在其他地区关于郊狼行为的记录进行对比分析。
牛群带来的直接益处
此前的研究表明,郊狼对牛群的捕食侵害在产犊高峰期最为严重(Boggess et al. 1978)。我们于2022年5月16日开始进行观察,此时正值产犊季节的尾声。在我们观察到的郊狼与牛群之间为数不多的直接互动(即郊狼接近某只幼犊,或冲向由母牛及其幼犊组成的群体)中,均发生在6月份——此时绝大多数幼犊均已出生(尽管我们在6月18日确实观察到了一次产犊过程,当时郊狼曾骚扰一头母牛以争夺胎盘)。我们研究区域内的大多数牧场主习惯将牛群安置在靠近自家宅邸的地方,直至4月或5月;因此,一旦牛群被转移至远离人类居所的区域,郊狼与带有幼犊的牛群发生互动的几率可能会随之增加。在两次观察中,母牛成功驱退了试图接近幼犊的郊狼;鉴于母牛在面对人类饲养员时也会表现出极强的护犊本能(Turner and Lawrence 2006),这一结果并不令人意外。不过,在我们观察到的唯一一次郊狼争食胎盘的事件中,尽管母牛曾试图追逐郊狼,但最终未能成功阻止它们将胎盘取走。由于缺乏不含幼犊的牛群样本,我们无法对“郊狼接近带有幼犊的牛群”与“郊狼接近不含幼犊的牛群”这两种情景下发生互动的几率进行对比分析。上述研究结果印证了此前的预测:即郊狼确实会利用牛群牧场来获取直接利益——具体方式包括捕食幼犊,或通过骚扰母牛来获取胎盘等副产品。然而,鉴于此类事件发生的频率极低,郊狼前往牛群牧场的主要动因似乎并非为了将活牛或其副产品作为直接的食物来源。相反,郊狼前往牛群牧场可能主要是为了捕食当地的野生猎物(详见“牛群带来的间接利益”一节)。此外,郊狼也可能选择在牛群附近徘徊,以便伺机寻找牛群中的虚弱个体进行捕食,同时顺便捕食该区域内的地松鼠。
郊狼还会对牛只尸体进行取食(即食腐),这对于身处牛群牧场环境下的郊狼而言,可能是一种重要的、周期性出现的食物资源。在本研究中,我们观察到郊狼的这种食腐行为发生频率较高;但这在一定程度上是由于我们在开展观察工作时,特意选择了那些已知存在牛只尸体的区域进行定点监测(详见“材料与方法”一节),从而导致数据结果可能存在一定的偏差。此前的研究已证实,郊狼为了获取牛只尸体作为食物,往往不惜长途跋涉(Kamler et al. 2004)。在我们观察到的两具牛只尸体周围,均有大量的郊狼频繁出没。成年母牛的死亡主要归因于分娩及天气相关的问题(APHIS 2010; Whiting et al. 2012),这表明春季可能会出现一波集中的牛尸资源。然而,郊狼对牛尸的取食行为在冬季可能更为普遍,因为此时郊狼及其他掠食者往往最依赖腐肉为生(Bekoff and Wells 1980; Morehouse and Boyce 2011)。在这一时节,牛尸通常会被堆放在靠近牧场居所的“堆骨场”内;与此同时,土拨鼠、地松鼠和昆虫等季节性食物来源也变得日益稀缺(Lingle et al. 2022)。如果大量郊狼在冬季乃至全年都将牛尸作为主要的食物来源,那么该地区的郊狼种群很可能正受益于牛尸这一资源的“补给”(Ciucci et al. 2020; Lingle et al. 2022);这种补给效应可能会导致郊狼在牧场区域实施更多的掠食行为,或引发其他滋扰问题(Goodale et al. 2015)。因此,建议就牛尸管理问题与当地牧场主进行磋商;因为若能通过将牲畜尸体运至场外处理或进行封闭式存放等手段来切断这种资源补给,将有助于降低人与郊狼发生冲突的风险(Beckmann and Berger 2003),并有助于控制郊狼种群的数量(Brunk et al. 2021)。在加拿大西部地区,省、畜牧生产者及当地社区之间已建立起合作伙伴关系,共同倡导将牲畜尸体管理作为一种有效的手段,旨在预防掠食者对牲畜的侵害,并避免掠食者与人类发生直接冲突。
牛群带来的间接益处
郊狼捕食本地猎物的频率远高于其与牛群互动的频率;然而,它们在牛群附近停留的时间却比预期要长,且在靠近牛群时频繁捕食地松鼠。这一发现印证了“间接益处假说”,即郊狼从牛群牧场中获益的方式,并不仅仅局限于将牛群本身作为食物来源。众所周知,在其他地区,郊狼也常在牛群附近频繁捕食小型哺乳动物(Fitch 1948);此外,理查森地松鼠(Richardson's ground squirrels)可能通过改善牧草质量,从而使放牧牛群从中受益(Newediuk et al. 2015)。牛群放牧会降低草的高度;而在植被较为低矮的区域,理查森地松鼠的分布往往更为普遍(Downey et al. 2006; Proulx et al. 2012),这可能归因于开阔视野有助于它们更有效地察觉捕食者。北极地松鼠(Urocitellus parryii (Richardson, 1825))在捕食者出没时,往往更倾向于选择视野开阔的栖息地;而在经历过一段高强度捕食压力的时期后,幸存下来的地松鼠也更倾向于利用此类视野良好的栖息地(Karels and Boonstra 1999)。在“草原国家公园”(Grasslands National Park)的高地栖息地中,理查森地松鼠在牛群放牧强度较高的区域分布更为密集(Bylo et al. 2014)。综合来看,上述研究表明:牛群对植被产生的影响,或许能使理查森地松鼠及其他高度依赖开阔栖息地的地松鼠物种从中受益。
关于“郊狼通过与牛群建立联系而获得间接益处——而非仅仅因为占据了牛群牧场(无论当时牛群是否在场)而获益”这一预测,我们的研究结果呈现出喜忧参半的印证效果。确实如预测所言,郊狼全天在牛群近处(距离小于100米)停留的时间,显著长于其在较远处停留的时间;且在这些近距离停留的时间中,有很大一部分是紧贴牛群(距离小于10米)度过的——这与“郊狼确实通过与牛群建立联系而获益”这一假设相吻合。郊狼捕食频率最高的猎物是理查森地松鼠;它们与牛群的伴随行为,可能恰好与地松鼠的活跃时段相吻合——地松鼠通常在日出后约一小时开始活跃,并在日落时分钻入洞穴后停止活动(Clark 1970)。然而,我们并未发现以下两种情境之间存在显著差异:即当郊狼紧贴牛群(近距离)时,其捕食地松鼠的频率或捕食成功率;与当牛群虽在同一区域内但距离较远时,郊狼的捕食表现。此外,在观察期间,那些身处牛牧场但周围无牛群,或牛群距离超过1000米的郊狼,其捕猎成功率与那些紧邻(距离小于100米)牛群的郊狼相似;尽管相比于无牛群在场或牛群距离超过1000米的情况,郊狼在靠近牛群时进行捕猎的频率往往更高。来自大黄石国家公园的证据表明,郊狼通常在低矮植被中捕食尤因塔地松鼠(Urocitellus armatus (Kennicott, 1863))(97%的捕猎行为发生在植被高度小于10厘米的环境中);且相比于高草丛或深雪环境,郊狼在短草地和浅雪环境中追捕田鼠(Microtus spp.)时的捕获成功率更高(Wells和Bekoff,1982)。本研究中观察到的郊狼与牛群之间的关联——而非仅仅是与“牛牧场”这一栖息地类型的关联——是否确实与捕食地松鼠的机会有关;以及若捕猎成功率确有提升,这种提升究竟归因于牛群活动所改造的栖息地中地松鼠密度较高,还是归因于郊狼在靠近牛群时所表现出的特定行为效应(例如利用牛群作为视觉或听觉掩护,以避免被地松鼠察觉),这些问题仍有待进一步深入研究。
鉴于我们的研究结果表明,郊狼从牛牧场中获益的方式并不仅仅局限于将牛群作为食物来源,我们推测郊狼在牛牧场中停留的时间应显著长于基于该类栖息地在景观中所占比例而得出的预期值(即相比于其他土地利用类型,郊狼对牛牧场表现出更高的利用偏好);然而,受限于研究条件,我们未能对这一推测进行实证检验。若能为研究对象佩戴无线电项圈,便可实现对个体郊狼跨栖息地的持续追踪——包括在夜间及清晨时段的追踪——从而进一步评估郊狼在行为上的差异,探究这些差异如何影响其与牛群的互动关系;同时也能借此确认郊狼是否会特意选择在一天中地松鼠最为活跃、从而最易被捕食的时段,前往牛牧场区域进行活动。
母牛及幼牛对郊狼的反应
我们的研究结果证实了此前的预测:当郊狼遭遇牛群时,母牛会表现出防御行为。在所有发生近距离接触(距离小于10米)的焦点观察案例中,有43%的案例显示母牛及幼牛对郊狼做出了防御性反应;其中包括郊狼两次直接逼近幼牛的事件。在这些防御反应中,有一例是当郊狼冲向母牛-幼牛混合群时,牛群紧密聚拢在一起;这种“聚拢”行为能够有效威慑郊狼,使其放弃对当地有蹄类动物的进一步追捕(Lingle 2001)。我们未发现防御行为与具体日期之间存在相关性的证据;这可能是由于某些特定时段的样本量过小,导致统计功效不足所致(图5)。因此,我们需要更大规模的样本数据来进一步确认以下问题:母牛是否在郊狼对幼牛构成最大威胁的时段表现出最强的防御性?母牛的抗捕食行为能否降低郊狼捕获幼牛的成功率?以及规模较大的郊狼群体是否更倾向于骚扰母牛或试图捕食幼牛?既往研究报告指出,定居型郊狼(因更易结成狼群)是大多数捕食事件的始作俑者(Sacks et al. 1999; Blejwas et al. 2002);且在捕食体型较大的猎物时,郊狼通常会采取群体协作的方式(Gese et al. 1988; Lingle 2000)。值得注意的是,在仅有的两次冲向母牛-幼牛混合群的事件中,发起冲锋的均为不止一只郊狼(分别由两只和五只组成的小群);此举很可能是为了惊扰牛群,试图将幼牛从群体中驱赶或分离出来。相比之下,那些直接逼近单个幼牛的郊狼均为独行个体。对此的一种合理解释是:郊狼群体主要负责实施主动捕猎行为,而独行的郊狼则更多地是基于机会主义心态去试探或侦察单个幼牛。
我们对牛牧场内郊狼行为的观察,有助于揭示可能影响牲畜被捕食风险的各类因素。我们的数据表明,郊狼利用牛牧场的主要目的是捕食当地的野生猎物,但它们也会伺机利用周期性出现的良机,对牛只尸体进行食腐,或试图捕食幼牛。因此,采取诸如阻止捕食者接触牲畜尸体之类的管理措施,对于制定行之有效的郊狼种群控制策略而言,可能具有至关重要的意义。
(致谢、参考文献:从略)
|
|